
7.Parametrized Sexy Globus Model with mate selection
The sensitivity analysis of the Global Model generates a new set of statistical simulation values for the evolution of intelligence. The description and the excellent results obtained are in the previous section about the Globus Model; now, there is an additional hypothesis of sexual selection to this model, which confirms the earlier results.
On the page on Evolution of the intelligence of the Conditional Evolution of Life book, the new Darwin-other experiment is explained –not yet carried out– to confirm the results of The EDI Study with a different methodology based on the effect on the evolution of the intelligence of the existing genetic mechanisms thanks to sexual differentiation.
The selection of a partner ** as an auxiliary mechanism of evolution has been a paradigm since the first development of the theory of evolution. Darwin himself wrote The Descent of Man and Selection in Relation to Sex (1871), introducing a new factor, mate selection or sexual selection, through which females or males choose those with the most attractive qualities as their partner.
Darwin was right about the choice of the most attractive qualities, but it is still another tautology, whatever those qualities are.
Intelligence is doubtlessly one of these desirable qualities for various reasons. However, from the viewpoint of the Sexy Globus model, it does not deal with imposing such a general hypothesis and much less given that the IQ of the mother and father are fixed.
When reflecting on the possibility of establishing another hypothesis to the Globus Model that could improve its general adjustment, the researchers thought of trying out the idea of relevance of the difference in intelligence between the father and the mother as a selection criterion or condition for the acceptation of the couple's configuration, in other words, mate selection.
It seems that intelligence is not a strict selection criterion when it comes to choosing a partner; but likewise, a noteworthy difference within the couple is not very common. Then, it should be possible to think of some optimization algorithms representing the indicated condition regarding sexual selection.
This small dissertation of evolutionary psychology about sexual selection becomes more complicated when thinking that there are two sets of intelligence corresponding to each one of the progenitors, and operating under diverse forms or conditions, as the Globus Model has already established. Perhaps the new hypothesis can go a little deeper into Darwin's statement about partner selection.
In short, the additional hypothesis introduced into the Sexy Globus model of mate selection is establishing the limit in the difference in intelligence that:
“The most potent chromosome of one of the couple's members has to be at least as powerful as the least potent chromosome of the other and vice versa.”
Here, the chromosome is a simplification to denote the part of intelligence inherited from one progenitor.

The psychological justification of the new hypothesis is that a person does not demand the same intelligence from others to like them, but to form a couple will demand that the other person can at least follow a conversation adequately. Just one chromosome can fulfill with the condition given that following the reasoning in a conversation does need certainty; in fact, the certainty derived from the Logical Verification of Information method comes within the initial argument, in other words, the two chromosomes of the person who talked first.
Although the explanation may not be very extensive, the relevant issue is that the Sexy Globus model substantially improves the adjustment due to the couple selection hypothesis. Nonetheless, the book of the Global Cognitive Theory deepens this effect.
The hypothesis will affect if introduced only the M2 or F2 chromosomes. Bearing in mind that the measured IQs collect the power of the significant or less forceful chromosome, the estimations of M2 and F2 will change considering the new information introduced in the model.
The Sexy Globus model somewhat improves with the individual variables (Otis test of fathers and mothers, Intelligence test Wechsler, and Stanford-Binet scales of children), but its response to selection criteria will be higher with centered variables. The GMCI with the rearrangement criteria M1F1° goes from 15.61 to 17 and the maximum r² from 0.89 to 0.97 for the objective function R° –see figures q063 and q077. For the objective function M & F, the GMCI is at 17.62 while it was 17.77, and the maximum r² rises from 0.89 to 0.97. The maximum values of r² usually correspond to the variable X6 – the average of six of the children's variables.
without mate selection
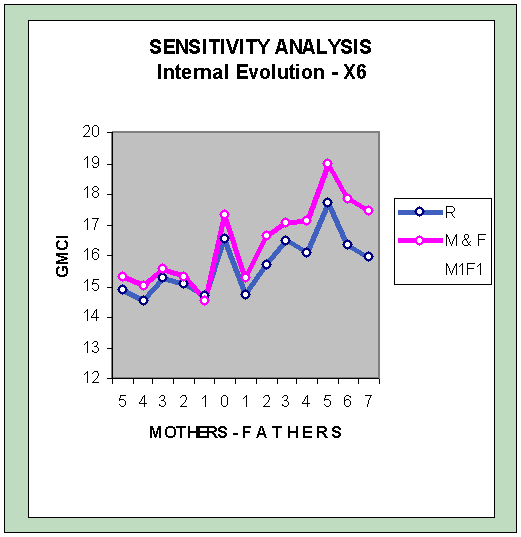
The sensitivity analysis carried out with the optimization algorithms and the variable X6 in the subsection of "Internal evolution" of the section "Simulation of the complexity of the real model" offers the following graph of the Globus Model (without sexual selection):
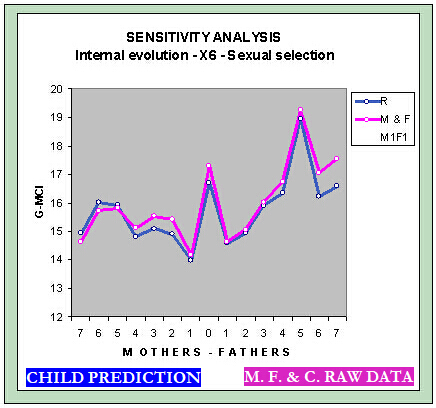
Repeating the same analysis performed with the variable X6 in the section of internal evolution of the Globus model, but with the additional hypothesis of partner selection, it gives the graph below of the Sexy Globus Model with the following aspects.
Of the graph's four peaks, three shifts upwards.
The correlations between the anticipated IQs for the objective function R° and the function M & F are more similar than before. The overall average of the function R° slightly rises, and the M & F slightly lower.
See the statistical table used for the sensitivity analysis of the intelligence evolution model, the internal evolution parameters with the additional hypothesis of sexual selection in terms of the minimum acceptable intelligence limit when forming a couple.
The interpretation is difficult, given the margin of the sensitivity of the introduced changes and the complexity of the Globus model. We have to take into account that only ten of the seventy IQs of the function R° have been affected in more than two percent of their value, but, trying to give a definite explanation of the mentioned aspects:
In respect to the first, it seems as though the model improves when its parameters are correct and that it worsens when the parameters are fictitious, which reinforces both the model and the introduced hypothesis.
In regards to the second, the function R° improves with additional information in its definition, while the function M & F, despite its peaks, decreases because it does not incorporate the effect of the hypothesis introduced on genes M2 and F2 given that it only collects information from M1 and F1 –that are the known IQs.
On the other hand, the Super Globus model’s results of the function R° improve as an objective function but not as rearrangement criteria. The reason might be that it deals with the average values of different possibilities because it incorporates the differences due to the genetic combination largely than the M1F1° criteria.
Globus Model with mate selection
The argument is similar to results of variable W°; in many cases, it has very high correlations but as rearrangement criteria, it usually is terrible because it incorporates the effects of genetic combination, of the functional limitations and affinity as well as the rest of the deviations due to the simulation of the errors of measurement and expression. Likewise, the children's variables tend to be outstanding rearrangement criteria because they do not incorporate the deviations due to genetic combination, affinity, internal evolution, or functional limitations.
If the hypothesis of sexual selection introduced were incorrect, the ICMG of the Sexy Globus Model could go down considerably, even for small changes. The sensitivity analysis graph shows this behavior when changing the internal evolution parameters for the variable X6, where one percent of potential increase by the fathers or mothers makes the GMCI drop drastically.
The Globus Model is just a graphical representation of the parameterization of the evolution in the Global Model, and the Sexy Globus Model refers to the introduction of the hypothesis of sexual selection.
In short, the proposed hypothesis of mate selection seems correct. The Sexy Globus model's coherence improves in general, and the correlations of the variables centered with the rearrangement criterion M1F1° increase substantially.